Preamble: In May 2018, HHMI Janelia hosted a wonderful conference on the evolution of neural circuits, organized by Albert Cardona, Melina Hale, and Gáspár Jékely. This is a transcript of a short talk I gave there. Caution: utter speculation!
Good evening. My name is Markus Meister. Many of you probably wonder what I am doing up here, and so do I. This started innocently enough when I wanted to attend this conference to learn about a different field. But the organizers asked for a poster abstract and I sheepishly complied. Then Albert Cardona unilaterally converted this poster to a talk, so now you know whom to blame. Let me also give two warnings. First, what I’ll present is pure speculation, there are no new results, only stringing together old results into questions. Fortunately it is timed to transition into the Apres-Ski part of the conference, so please consider this as part of the evening entertainment. Second I want to deliver a trigger warning. For those of you not at an American university, we professors are encouraged to warn our sensitive students when a lecture threatens to bring up a subject they might find offensive. The present talk will include notions reminiscent of Jean-Baptiste Lamarck, in particular his idea that acquired characters are inheritable.

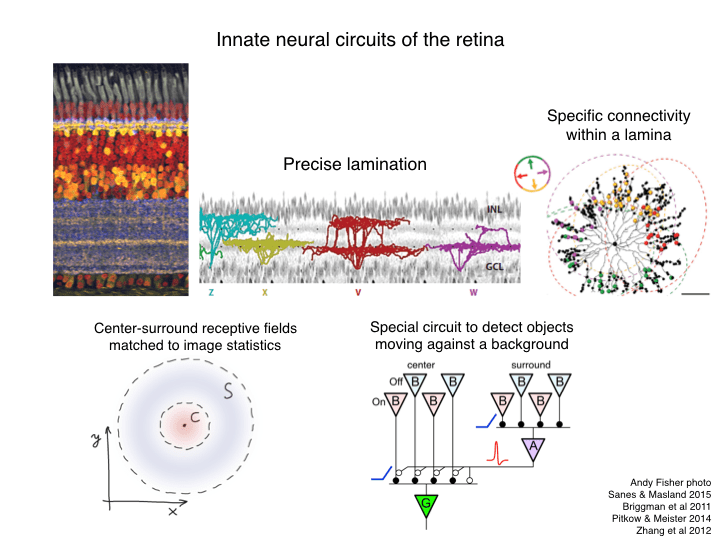
I want to consider how neural circuits evolved to what we find today. In any given brain one finds some circuits that are essentially specified by the genome. A good example of that is the retina: pretty much the same circuit in every member of the species, with rather little influence from visual experience. It’s a rather complex and precise network among 80 or so cell types. That ultimately produces about 30 different types of retinal ganglion cell with different visual functions. Those functions are exquisitely well adapted to the visual environment. For example the well-known center-surround organization of receptive fields matches beautifully with the statistical structure of natural scenes to produce a kind of image compression that functions a bit like JPEG. Delving in more detail we find a type of retinal ganglion cell designed to pick out a moving object against a moving background, a function that is obviously useful for both predator and prey animals. Each of these circuits relies on precise connectivity among a dozen or so different cell types. And that connectivity is ultimately determined by the developmental program stored in the genome.
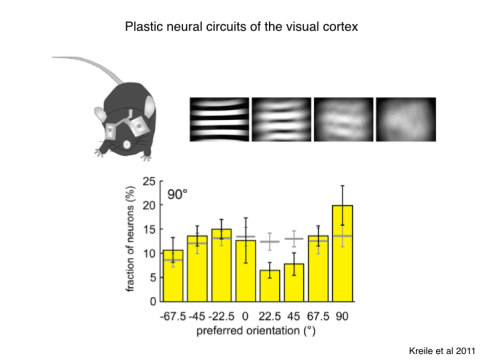
The common explanation for how this adapted circuit came about is through the cumulative effects of mutation at the gene level and natural selection at the level of behavior. And I’m sure that is a large part of the story. But we have to consider that the brain also has powerful mechanisms for adapting in real time. In the visual system, if we just take a couple steps forward from the retina, we find circuits in the mammalian cortex that are much more dependent on environmental exposure, and can adapt plastically to many different environments. For example you can raise a mouse with cylindrical goggles so it experiences almost exclusively patterns with one orientation. After just a few weeks of this the cortex has reorganized to preferentially encode that orientation. The detailed synaptic circuits that accomplish this are acquired through experience, shaped by various cellular mechanisms of learning. In the course of a lifetime an organism acquires a great deal of experience from the environment that gets stored in synaptic connections within these plastic circuits. But the common notion of brain evolution says that all this experience is lost to the next generation.
So I would like to revisit this question now: Is it possible that some of the acquired knowledge about the environment is in fact passed on genetically to the offspring. And more specifically, is it possible that the innate neural circuits we find today are really the accumulated result of many generations of acquired circuitry that eventually got frozen in the genome. This of course relates to Lamarck’s doctrine that organisms can pass acquired traits on to their offspring. Although it is worth remembering that Darwin also speculated about this possibility. But why revisit this question now? Do we know of any example where an organism learns from the environment and stores that knowledge in the genome for the benefit of future generations?
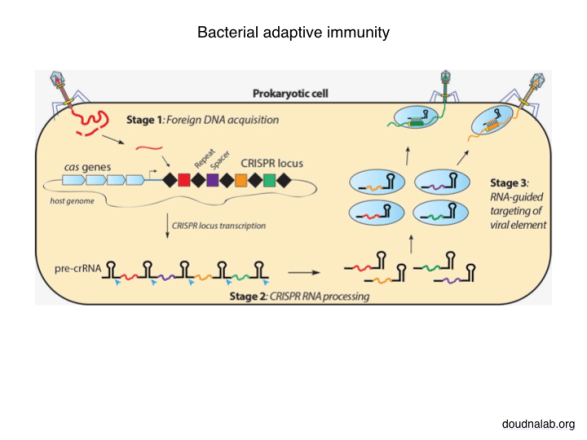
Of course, you will say: the ancient bacterial immune system. By now even theoretical physicists have been caught up in the hoopla around CRISPR. But beyond the technological promise of gene editing and the associated patent fights, I find the biological function of this system more fascinating. The molecular machinery in the bacterium identifies foreign DNA, like from a virus infection, chops it into small pieces, and inserts those pieces at a special location in the bacterial genome. Meanwhile other proteins read the sequences stored in that memory and look for matching items floating around in the cell to destroy them. The descendants of this cell inherit the stored memories and are therefore better prepared for threats in the environment. Of course this clever mechanism itself had to evolve somehow, but once in place it opens the door to a Lamarckian mechanism of inheritance.
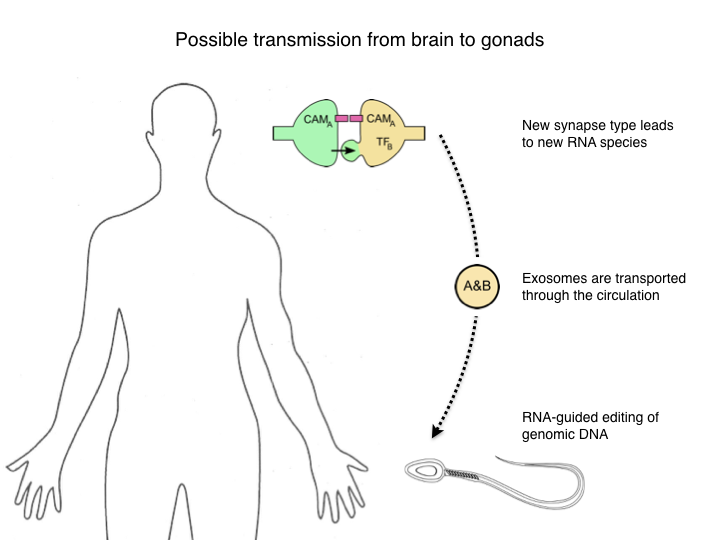
Given that some early forms of life have developed a way to record experience in the genome, it seems plausible that Nature has continued to use this powerful principle. Now there are two aspects of this bacterial system that make it particularly easy to transmit acquired experience genetically. First, the information gathered about the environment is already in the form of genetic sequences, such as a viral genome. Second, the cell that is sampling the environment is the same cell that transmits the genome to the descendants. In multi-cellular animals of course the germ line is separated from the rest of the body, and the environmental experience is not directly available as a DNA sequence. So in thinking about how this might work in animals we need to try and solve both problems: Turn synaptic connectivity into sequence information and then transmit that from the brain to the gonads.
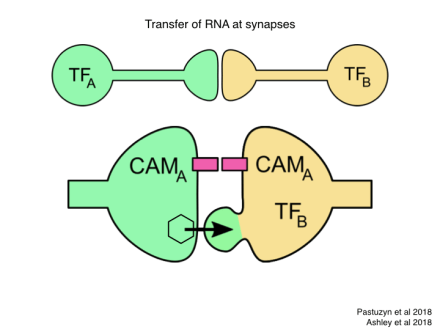
So let’s think about a concrete example with minimal complexity, what is commonly called a “toy model”. We suppose that lifetime experience has set up new synaptic circuits in the brain. This leads to new synapses between two cell types, call them A and B. Each of these types is defined by expression of a specific transcription factor, TF-A and TF-B. We would like to communicate this event to the germline in such a way that in the next generation these two neurons are more likely to form a synapse all by themselves, without the need for learning and plasticity. Of course we don’t require that this happens deterministically after just one generation. Even if the probability of forming such a connection is enhanced just by a tiny amount, that biases the genetic program for this neural circuit a little bit, and that bias can accumulate over generations.
So how do we imagine that information about this event in the brain gets to the germ cells? Whatever carries that signal has to be a information-rich message, because the event is rather specific. Assuming the brain has about 1000 cell types, and we want to encode synapses between any two of them, that requires abut a million different symbols. So the signal has to be something with a million possible values. Obviously we’re not talking about some hormone. Instead the likely carrier would be a nucleotide sequence, like RNA. Short RNAs would be sufficient, since 10 bases can already encode a million signals.
Now in recent years there has been a flurry of discoveries about RNA mechanisms, and I want to point to three of them that may be relevant for the scheme we’re considering here. Starting with synapse formation: We learned just a few months ago that at a synaptic junction cells can pass RNA to each other inside a virus-like capsid, and that alters gene expression in the partner neuron. One might imagine that this could be used to strengthen a synaptic connection. For example if cell A expresses a homotypic cell adhesion molecule CAM-A, the mRNA for that could transfer to cell B, leading to expression of CAM-A in that neuron as well and consequent cell adhesion. More importantly, cell B now has a complement of RNAs that it did not have before this event, for example the mRNAs for transcription factor TF-B and for the adhesion molecule CAM-A. That can lead to the production of new species of RNA that did not exist before.
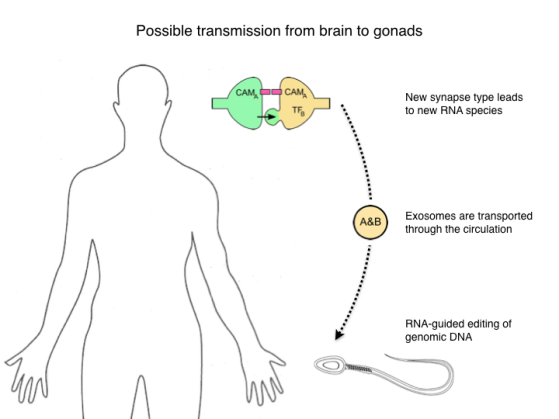
Second, how can that information flow to the germ line? One such mechanism are extracellular vesicles, sometimes called exosomes (although that term is ambiguous). These are small vesicles extruded from the plasma membrane of many cells. They can carry protein, RNA, and DNA. They provide a route for cells to exchange all these information-rich signals. They are transported through the blood and remarkably enough they can cross the blood-brain-barrier. Separate evidence says they are taken up in the testes. So in principle a route exists for RNA molecules to convey information about such peripheral events to the germline.
Finally, how could that information be used to alter the genome? Again here we are learning about new mechanisms that are potentially relevant. In particular there are a number of reports of DNA editing guided by sequences in non-coding RNA molecules. Of course the man-made systems based on CRISPR-Cas9 present just such a mechanism, but there seem to be related endogenous processes as well.
How would one test whether something like this is afoot? There have been a good number of studies of transgenerational learning, starting perhaps with the infamous experiments on cannibalistic planarians of the 1960s. I don’t think that’s a promising route, because any bias for directed evolution from such a cascade would likely be too weak to be observed in one or a few generations. Instead one might look for some essential mechanistic ingredient, like the postulated transport of polynucleotide signals from brain to germ cells. Is it possible, for example, to chemically label all the RNA synthesized in the brain and look for species that make their way to the testes?
As I said at the outset, these ruminations are purely speculative. But it seems to me there is still room for discovery in the ample dark regions of the genome, and some mechanism of this kind would perhaps fit there. If so it could fundamentally change our view of how the complex circuits of the brain came about. In any case, I hope that some of the napkins at the bar get used to sketch some candidate mechanisms or even to prove why it cannot possibly work.
Literature cited on the slides:
Ashley J, Cordy B, Lucia D, Fradkin LG, Budnik V, Thomson T. Retrovirus-like Gag Protein Arc1 Binds RNA and Traffics across Synaptic Boutons. Cell. 2018;172: 262-274.
Briggman KL, Helmstaedter M, Denk W. Wiring specificity in the direction-selectivity circuit of the retina. Nature. 2011;471: 183–188.
Kreile AK, Bonhoeffer T, Hübener M. Altered visual experience induces instructive changes of orientation preference in mouse visual cortex. J Neurosci. 2011;31: 13911–20.
Pastuzyn ED, Day CE, Kearns RB, Kyrke-Smith M, Taibi AV, McCormick J, et al. The Neuronal Gene Arc Encodes a Repurposed Retrotransposon Gag Protein that Mediates Intercellular RNA Transfer. Cell. 2018;172: 275-288.
Pitkow X, Meister M. Neural computation in sensory systems. In: Gazzaniga MS, Mangun GR, editors. The Cognitive Neurosciences. 5th ed. Cambridge, MA: MIT Press; 2014. pp. 305–318.
Sanes JR, Masland RH. The types of retinal ganglion cells: current status and implications for neuronal classification. Annu Rev Neurosci. 2015;38: 221–246.
Zhang Y, Kim IJ, Sanes JR, Meister M. The most numerous ganglion cell type of the mouse retina is a selective feature detector. Proc Natl Acad Sci U S A. 2012;109: E2391-8.
